
Exosome secretion from hypoxic cancer cells reshapes the tumor microenvironment and mediates drug resistance

 ,
, Abstract
Hypoxia is a common phenomenon in solid tumors as the poorly organized tumor vasculature cannot fulfill the increasing oxygen demand of rapidly expanding tumors. Under hypoxia, tumor cells reshape their microenvironment to sustain survival, promote metastasis, and develop resistance to therapy. Exosomes are extracellular vesicles secreted by most eukaryotic cells, including tumor cells. They are enriched with a selective collection of nucleic acids and proteins from the originating cells to mediate cell-to-cell communication. Accumulating evidence suggests that exosomes derived from tumor cells play critical roles in modulating the tumor microenvironment (TME). Hypoxia is known to stimulate the secretion of exosomes from tumor cells, thereby promoting intercellular communication of hypoxic tumors with the surrounding stromal tissues. Exosome-mediated signaling pathways under hypoxic conditions have been reported to cause angiogenesis, invasion, metastasis, drug resistance, and immune escape. Recently, the programmed cell death ligand-1 (PD-L1) has been reported to reside as a transmembrane protein in tumor exosomes. Exosomal PD-L1 was shown to suppress T cell effector function in the TME and cause drug resistance to immune checkpoint therapy. This review provides an update about the pivotal role of tumor-derived exosomes in drug resistance to chemotherapy and immunotherapy, particularly under hypoxic conditions. Emerging strategies that target the exosomes in the hypoxic TME to enhance the antitumor efficacy are discussed.
Keywords
INTRODUCTION
Hypoxia is a well-known hallmark of solid tumors when the tumor vasculature cannot provide adequate oxygen to support the aggressive growth of rapidly expanding tumors. Preclinical studies demonstrated that hypoxia mediates resistance to various modalities of cancer therapy, including chemotherapy, radiotherapy, and immunotherapy[1,2]. Tumor hypoxia may also promote invasion and metastasis. Extensive evidence is also available from clinical investigations to suggest that highly hypoxic tumors are associated with treatment failure, increased incidence of distant metastases, and dismal disease-free and overall survival[3].
Exosomes are a unique form of extracellular vesicles with endosomal origin and sizes ranging from 30 to 100 nm. They are secreted from diverse cell types upon the fusion of multivesicular bodies with the plasma membranes[4]. Exosomes mediate intercellular crosstalk by transferring mRNAs, microRNAs (miRNAs), and proteins from donor to recipient cells[5,6]. Cargoes loaded in exosomes are biologically active when taken up by the recipient cell, and they lead to various downstream functions[7]. Tumor-derived exosomes have been shown to facilitate the intercellular transfer of pro-tumorigenic factors in the tumor microenvironment (TME)[8,9]. They promote angiogenesis, invasion, and proliferation in recipient cells to support tumor growth and a pro-metastatic phenotype. In a recent proteome profiling study of exosomes derived from human primary and metastatic colorectal cancer cells, selective enrichment of metastatic factors and signaling pathway components was observed[10]. In glioma, exosomes have been reported to convey signals between the tumor and TME to facilitate bidirectional communication[11]. Hypoxia is known to stimulate the secretion of exosomes from tumor cells, thereby promoting cell-to-cell communication between the hypoxic tumors and the surrounding stromal tissues. Exosomal cargoes are also altered under hypoxic conditions to stimulate angiogenesis, invasion, metastasis, therapeutic resistance, and immune escape[12].
In this article, we summarize the critical role played by hypoxic tumor-derived exosomes in tumor progression and resistance to cancer therapy. Novel approaches to target the exosomes in the hypoxic TME to potentiate the anticancer efficacy are discussed.
HYPOXIC TME
TME is a dynamic and complex system around a tumor, which is composed of blood vessels, fibroblasts, extracellular matrix fibers, immune cells, and signaling molecules, supporting proliferation, metastasis, and therapy resistance of tumor cells[13]. The rapid proliferation of cancer cells and aberrant blood vessel formation create hypoxic conditions in malignant tumors. Hypoxia is considered a hallmark of TME, which is generally defined by a low oxygen tension of below 10 mmHg[14]. It is also known to orchestrate the various malignant phenotypes of cancer cells by activating multiple oncogenic signaling pathways. At the molecular level, the hypoxic TME is largely regulated by the hypoxia-inducible factor (HIF) family of transcriptional factors[15]. Accumulating evidence suggests that tumor-derived exosomes play a critical role in invigorating the interaction between cancerous and non-cancerous cells in the hypoxic TME to propel cancer progression[16].
EFFECT OF HYPOXIA ON TUMOR-DERIVED EXOSOME
Induction of exosome release
TME-associated cells have been shown to secrete more exosomes than normal cells to promote intercellular communication and nutrient exchange[17]. In the clinical setting, the number of circulating exosomes collected from the blood of cancer patients was estimated to be more than two-fold higher than that of healthy subjects[18]. Granulocytic myeloid-derived suppressor cells (G-MDSCs) in tumors were reported to produce more exosomes than those found in the spleen[19]. Indeed, more secretion of exosomes from cancer cells has been reported in breast cancer[20], colorectal cancer[21], gastric cancer[22], glioma[23], hepatocellular carcinoma[24], and pancreatic cancer[25]. Interestingly and in contrast, hyperoxia (excessive oxygen tension) has been reported to reduce the number of exosomes released from colorectal cancer cells[19]. It is noteworthy that the induction of exosome secretion in hypoxia may be universal because more exosomes were also released from non-cancerous cells (including bone, cardiac, and kidney cells) under hypoxic conditions[26-28]. To date, the detailed mechanism leading to increased exosome release in hypoxia is still not clear. Nevertheless, two Rab GTPases (Rab27a and Rab27b) were shown to promote exosome secretion by facilitating the docking of the multivesicular bodies at the plasma membrane[29], whereas Rab7 was reported to direct exosomes to lysosomes for degradation[30]. In ovarian cancer cells, hypoxia was shown to upregulate Rab27a but downregulate Rab7 to promote exosome release[31].
Increase in exosomal heterogeneity
Exosomes secreted from different cells contain different cargoes and markers. On the other hand, exosomes released from the same cell line can carry different constituents[18]. While this heterogeneity is advantageous for exosome applications, it poses obstacles to a thorough understanding of exosome biology[32,33]. Depending on the cell state, exosomes of different sizes and carrying different cargoes are secreted[34]. To this end, exosomes secreted from glioma cells were shown to reflect the hypoxic status, and they mediate hypoxia-dependent activation of neovascularization during tumor development[34]. The heterogeneous exosome population could have a different functional effect on the recipient cells[35,36].
Production of smaller exosome
The size of exosomes released from mammalian cells is known to vary considerably in individual cells because of their unique exosome biogenesis process[36]. Interestingly, it has been reported that exosome size is associated with different disease states. In non-small cell lung cancer (NSCLC) patients, a smaller exosome detected in the pulmonary vein is associated with a shorter time to relapse and shorter overall survival after curative surgery[37]. It is generally believed that hypoxia tends to release smaller exosomes. Relatively smaller exosomes have been reported in different cancer cell lines, including colon[21], pancreatic[25], and prostate[38] cancer cells. Under hypoxic conditions, it is believed that the supply of cellular materials for membrane synthesis may not meet the demand for exosome production, thus leading to the secretion of smaller exosomes[12]. It is also hypothesized that smaller exosomes could be transmitted more easily via the blood circulation to metastatic sites to form the pre-metastatic niches because hypoxia changes the hemodynamics in the tumor vasculature[12]. Moreover, smaller exosomes could be internalized faster and more efficiently than larger exosomes in the recipient cells[39]. An ongoing prospective clinical trial (NCT02310451) is underway, which examines exosome size and various other parameters as potential biomarkers in melanoma patients.
Influence on exosomal cargo sorting
It is known that exosome content is not simply reflecting the cellular content of its origin. Some sorting mechanisms are in place to facilitate specific cargo sorting processes. Hypoxia has been shown to affect the sorting of the following three major cargoes (i.e., nucleic acids, proteins, and lipids) into exosomes.
Nucleic acids
The effect of hypoxia on the nucleic acid content in tumor-derived exosomes has recently been summarized in a few excellent reviews[40,41]. It is noteworthy that only limited studies have reported the effect of hypoxia on cancer-derived exosomal DNA. On the other hand, the alteration and biological effects of non-coding RNAs (ncRNAs) in tumor-secreted exosomes under hypoxia have been extensively studied. Representative examples are summarized in Table 1. Various mechanisms have been proposed to regulate the expression of ncRNAs in hypoxia. For the HIF-dependent mechanism, the binding of HIF-1α and/or HIF-2α to the hypoxic response element (HRE) of the gene promoter of miR-155 has been reported[42,43]. Moreover, miRNAs are differentially sorted into exosomes according to their specific sequence or motif[44]. A few RNA binding proteins, such as SYNCRIP, were shown to direct specific miRNAs sharing a common extra-seed sequence hEXO motif for enrichment in the exosomes[45]. Moreover, hypoxia also affects RNA alternative splicing[46,47] and RNA editing[48], which could make specific miRNAs more suitable for loading into tumor-derived exosomes. On the other hand, hypoxia was also reported to regulate a few facilitators which load nucleic acids into exosomes. The RNA binding proteins YBX1 and hnRNPA1 were reported to mediate the selective loading of miR-133 and miR-1246, respectively, into tumor-derived exosomes under hypoxia[49,50].
Representative non-coding RNA cargoes secreted in hypoxic cancer cell-derived exosomes to modulate the tumor microenvironment
| Non-coding RNA | Donor cell | Recipient cell | Biological function (mechanism) | Reference |
| let7a | Melanoma | Macrophage | Induce M2 polarization of TAM and promote oxidative phosphorylation to support cancer growth (downregulation of insulin-AKT-mTOR signaling) | [121] |
| linc-RoR | Hepatocellular carcinoma | Hepatocellular carcinoma | Promote cancer cell proliferation; mediate chemoresistance (induction of PDK1 and HIF-1α protein expression by suppressing miR-145) | [78] |
| lncRNA UCA1 | Bladder cancer | Bladder cancer | Promote cancer growth; stimulate migration and invasion (induction of EMT) | [95] |
| miR-10a | Glioma | MDSC | Activate MDSC (regulation of RORA/IκBα/NF-κB signaling) | [113] |
| miR-21 | Glioma | MDSC | Activate MDSC (regulation of PTEN/PI3K/AKT signaling) | [113] |
| miR-23a | Lung cancer | Endothelial cells | Promote angiogenesis; increase vascular permeability (inhibition of the propyl hydroxylases PHD1 and PHD2, thereby stabilizing HIF-1α protein; inhibition of the tight junction protein ZO-1) | [92] |
| miR-24-3p | Nasopharyngeal carcinoma | T cell | Inhibit T cell proliferation and differentiation (inhibition of FGF11; upregulation of p-ERK, p-STAT1, and p-STAT3) | [103] |
| miR-25-3p | Breast cancer | Breast cancer and macrophage | Promote cell proliferation and migration (stimulation of IL-6 secretion from macrophage via NF-κB signaling) | [94] |
| miR-125b-5p | Ovarian cancer | Macrophage | Induce M2 polarization (regulation of SOCS4/5-STAT3 pathway) | [120] |
| miR-210 | BMSC | Lung cancer | Promote metastasis (induction of STAT3 to mediate EMT) | [73] |
| miR-301a-3p | Pancreatic cancer | Macrophage | Induce M2 polarization (downregulation of PTEN and activation of PI3K/Akt signaling) | [118] |
| miR-5100 | BMSC | Lung cancer | Promote metastasis (induction of STAT3 to mediate EMT) | [97] |
Hypoxia has been shown to regulate the expression of several ncRNAs, which affect the expression of HIF-1α and subsequently form positive or negative feedback loops to modulate the hypoxic TME[51,52]. In hypoxic gastric cancer cells, such a positive feedback loop involving miR-301a-3p, PHD3, and HIF-1α has been reported[53]. miR-301a-3p was upregulated in hypoxic gastric cancer cells and the tumor-secreted exosomes[53]. miR-301a-3p was subsequently shown to suppress the hydroxylase PHD3 and thus promote the protein stability of HIF-1α to maintain the hypoxic response. In pancreatic cancer, the hypoxic induction of a HIF-1α-stabilizing circular RNA (cirZNF91) in cancer-secreted exosomes was reported to promote chemoresistance in normoxic cancer cells[54]. Upon transmission to normoxic cells, circZNF91 was found to bind competitively to miR-23b-3p, subsequently abolishing the inhibition of miR-23b-3p on its target Sirtuin1 (SIRT1). The upregulated SIRT1 was shown to increase the deacetylation-dependent stability of HIF-1α protein, thus promoting glycolysis and chemoresistance in the recipient normoxic cancer cells[54].
Proteins
Similar to nucleic acids, proteins loaded into tumor-derived exosomes are not necessarily proportional to the cellular protein composition[55]. However, the precise mechanism contributing to specific exosomal protein sorting remains obscure. It has been postulated that hypoxia could influence the protein loading process of tumor-derived exosomes by regulating ubiquitination. Ubiquitinated proteins are recognized by ubiquitin-binding domains within multivesicular endosomes for degradation, thus limiting the amount of membrane available for exosome formation[56]. To this end, hypoxia is commonly known to control protein ubiquitination and ubiquitination-associated enzymes[57].
Cell membrane glycoproteins are also known to participate in cell-to-cell and cell-environment communication[58]. Interestingly, the expression profile of glycoproteins in tumor-derived exosomes is different from that of healthy cells[59]. Moreover, glycoprotein expression could be affected by hypoxia[60]. As glycans are known to regulate protein sorting and uptake into exosomes, hypoxic cells have been shown to take up more exosomes in a proteoglycan-dependent manner[61].
Proteins secreted in tumor exosomes are also known to participate in hypoxia-associated responses. In breast cancer, metastasis-associated protein 1-loaded exosomes were reported to transfer in between cancer cells to regulate the response to hypoxia and estrogen signaling[62]. Taken together, hypoxia promotes numerous malignant phenotypes of cancer cells by altering the exosome protein heterogeneity, whereas proteins in tumor-derived exosomes could contribute to the hypoxic response. Table 2 summarizes the representative exosomal protein cargoes and other constituents that are preferentially secreted from hypoxic tumors to module the TME.
Exosomal protein cargoes and other constituents are preferentially secreted from hypoxic tumors to modulate the tumor microenvironment
| Cancer type | Type of cargo | Biological function and mechanism | Reference |
| Colorectal cancer | Wnt4 protein | • Intercellular communication with normoxic cancer cells • Exosomal Wnt4 promoted the translocation of β-catenin to the nucleus in normoxic cells • Activation of β-catenin signaling enhanced cancer cell motility and invasion | [137] |
| Glioblastoma | TSP1, VEGF, LOX protein | • Promote cancer growth, angiogenesis, and metastasis | [138] |
| Glioblastoma | MMPs, IL-8, caveolin 1, PDGFs, and lysyl oxidase | • Induce angiogenesis in vitro and ex vivo through phenotypic modulation of endothelial cells • Stimulate endothelial cells to secrete potent growth factors and cytokines and to activate pericyte PI3K/Akt signaling to promote migration | [34] |
| Lung cancer | TGF-β1 | • TGF-β1 downregulates NKG2D on the cell surface of NK cells to suppress NK cell cytotoxicity | [116] |
| Nasopharyngeal carcinoma | MMP13 | • Promote migration and invasion | [139] |
| Nasopharyngeal carcinoma | HIF-1α | • Promote EMT to induce migration and invasion | [140] |
| Prostate cancer | Tetraspanins (CD63 and CD81), heat shock proteins (HSP90 and HSP70), and Annexin II | • Remodel the epithelial adherens junction pathway to enhance invasiveness and stemness of naïve prostate cancer cells | [38] |
| Prostate cancer | Lactic acid | • Under chronic hypoxia, prostate cancer cells secrete more exosomes as a survival mechanism to remove metabolic waste | [141] |
Lipids
Various lipid components are important building blocks for exosome membranes. They include phosphatidylcholine, phosphatidylethanolamine, phosphatidylinositol, phosphatidylserine, phosphatidic acid, cholesterol, ceramide, sphingomyelin, and glycosphingolipids[59]. They play critical roles in the biogenesis of exosomes. Sphingomyelin is hydrolyzed to ceramide, which promotes the budding of multivesicular bodies from the endosomes[63]. On the other hand, ceramide is metabolized to sphingosine 1-phosphate, which interacts with the inhibitory G protein-coupled receptors to induce exosome biogenesis[64]. Interestingly, knockdown of a key cholesterol lipid efflux transporter ABCG1 was found to inhibit cancer growth, concomitantly with the intracellular accumulation of exosomes and the exosomal cargo[65]. Phosphatidylserine has been reported to play a critical role in facilitating the uptake of exosomes secreted from hypoxia-induced stem cells by human umbilical cord endothelial cells[66]. Under hypoxic conditions, triglyceride was found to accumulate in prostate cancer cells and the secreted exosomes. Importantly, these exosomes were shown to promote cancer proliferation and invasion following reoxygenation[67]. Hypoxia is known to upregulate ceramide expression, which is proposed to promote the exosome release[68].
Effect on exosome uptake by recipient cells
Little is known about the effect of hypoxia on the intercellular transfer and uptake of tumor-derived exosomes from other components within the TME. A hypoxic environment is known to promote glycolysis and lactic acid production. Excess production of lactic acid results in acidic pH in the hypoxic TME. Given that an acidic environment is more suitable for the stability and isolation of exosomes[69], the intracellular transport of exosomes may benefit from a hypoxic and acidic TME. It has also been proposed that the uptake of cancer-secreted exosomes by the recipient cells was found to be more efficient under hypoxia[70]. The smaller exosomes secreted from hypoxic tumors, as described above, may facilitate the more efficient intercellular transfer. However, this hypothesis has not been verified by a detailed experimental investigation. On the other hand, hypoxic tumor cells are also known to take up more exosomes from the surroundings[61]. Upon the recognition and binding of exosomes to recipient cells, the former is internalized by various processes, including endocytosis via clathrin, caveolae, or lipid raft-dependent manner. Under hypoxic conditions and the closely associated acidic cellular environment, exosome uptake could be promoted by lipid raft-dependent endocytosis[61], caveolin-dependent endocytosis[71], and phagocytosis[72].
REGULATION OF TME BY HYPOXIC TUMOR-DERIVED EXOSOMES
As described in the previous section, hypoxia significantly alters the properties of exosomes secreted from cancer cells. Accumulating evidence indicates that the altered tumor exosomes are responsible for the reshaping of TME, thereby promoting cancer cell proliferation, chemoresistance, metastasis, and angiogenesis [Figure 1].
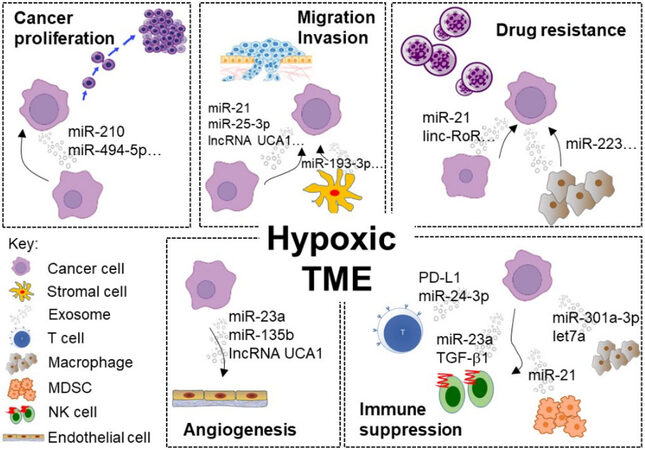
Figure 1. Representative examples of exosome-mediated intercellular communication within the hypoxic TME driving cancer progression, chemoresistance, and immune suppression. (1) Cancer proliferation: Hypoxic cancer-secreted exosomes were enriched with miRNAs supporting cell survival (e.g., miR-210) in the neighboring cancer cells. (2) Drug resistance: Numerous ncRNAs (e.g., linc-RoR and miR-21) were transferred via exosomes from hypoxic and resistant cancer cells to sensitive cells and induced drug resistance. (3) Migration and invasion: exosomes containing various ncRNAs (including lncRNA UCA1 and miR-193-3p) facilitate cancer–cancer or cancer–stromal intercellular communication to stimulate migration and invasion by modulating EMT. (4) Angiogenesis: ncRNAs (including miR-23a and lncRNA UCA1) were enriched in the exosomes secreted from hypoxic tumor cells to promote tumor vascular endothelial cell proliferation and angiogenesis in HIF-1α-dependent or -independent pathway. (5) Immune suppression: Exosomes enriched with miRNAs (e.g., miR-23a and let-7a) and other immunosuppressive molecules (e.g., PD-L1 and TGF-β1) were secreted from hypoxic tumors to promote an immunosuppressive TME. TME: Tumor microenvironment.
Promotion of cancer proliferation and chemoresistance by hypoxic tumor-derived exosomes
Within the hypoxic TME, cancer cells are known to secrete pro-tumorigenic molecules in the exosomes to promote cancer survival and proliferation. miR-210 is one of the most extensively studied hypoxia-induced miRNAs driving cancer progression[73]. In breast cancer, the abundance of miR-210 was reported to be remarkably higher in the exosomes derived from hypoxic cancer cells than those from normoxic ones, which allows the cells to sustain survival under hypoxia[74]. A set of differentially expressed exosomal miRNAs has been identified in the exosomes secreted from patient-derived melanoma cells under hypoxic culture conditions[75]. Hypoxia was found to upregulate miR-494-5p, miR-4497, miR-513a-5p, and miR-6087 but downregulate miR-125b-5p, miR-21-5p, and miR-3934-5p in the exosome[75]. Interestingly, the alteration of miRNAs was closely associated with cancer survival according to bioinformatics pathway analysis[75]. Exosomes secreted by human glioma were also shown to promote the differentiation of neural stem cells into astrocytes[76]. Transcripts related to cell proliferation and astrocyte differentiation were found to be remarkably upregulated in human mesenchymal stem cells when co-cultured with glioma-secreted exosomes[76]. Hypoxic tumor-secreted exosomes may represent an important therapeutic target that mediates the aggressiveness of glioma.
Hypoxia is known to mediate chemoresistance by regulating the cell cycle, autophagy, cell senescence, and drug efflux transporters. In recent years, the emerging role of hypoxic cancer cell-derived exosomes in reshaping the TME and causing chemoresistance is also revealed. In NSCLC, hypoxic cancer-derived exosomes have been shown to induce cisplatin resistance in normoxic cancer cells through the transmission of miR-21[77]. The transfer of miR-21 from hypoxic cell-derived exosome to normoxic cancer cells was demonstrated to downregulate PTEN and the PI3K/Akt pathway, which subsequently induced cisplatin resistance[77]. In hepatocellular carcinoma, the abundance of a stress-responsive lncRNA (linc-RoR) was significantly increased in hypoxic cancer-derived exosomes than in their normoxic counterpart[78], which is associated with resistance to sorafenib and doxorubicin. Linc-RoR was shown to induce TGF-β, thereby suppressing chemotherapy-induced cell death but promoting tumor-initiating cell proliferation[78]. Stromal cells, such as cancer-associated fibroblasts (CAFs) in the TME, could also mediate chemoresistance in cancer cells. miR-223 was upregulated in TAMs and TAM-derived exosomes under hypoxia[79]. miR-223 loaded in hypoxic exosomes was shown to reduce apoptosis and induce drug resistance in ovarian cancer by downregulating PTEN and thus activating PI3K/Akt signaling[79].
Moreover, exosomes have also been shown to mediate the transfer of the drug-resistant phenotype. Drug-sensitive cancer cells have been shown to become drug-resistant following the incorporation of exosomes shed from drug-resistant cancer cells[80-84]. Moreover, exosomes were shown to be involved in the intercellular transfer of functional ABCB1 (P-gp) from multidrug-resistant donor cells to drug-sensitive recipient cells[81,85-87]. Furthermore, exosomes have also been reported to mediate drug resistance by exporting specific drugs via the exosome pathway[88] and neutralizing antibody-based chemotherapy[89].
Induction of cancer angiogenesis by hypoxic tumor-derived exosomes
The induction of angiogenesis by hypoxia has been extensively studied[90]. More recently, accumulating evidence demonstrates that hypoxic tumor-derived exosomes played a significant role in angiogenesis. In malignant glioblastoma multiforme, exosomes derived from cancer cells under hypoxia were shown to induce angiogenesis by stimulating cytokine and growth factor secretion from endothelial cells, subsequently promoting pericyte migration[34]. In pancreatic cancer, the exosomal lncRNA UCA1 secreted from cancer cells under hypoxic conditions was shown to promote angiogenesis via a miR-96-5p/AMOTL2/ERK1/2 pathway[91]. In lung cancer, miR-23a secreted in tumor-derived exosomes in hypoxia was reported to target the key HIF-1α regulators (propyl hydroxylases PHD1 and PHD2), thereby sustaining the overexpression of HIF-1α and promoting angiogenesis[92]. Moreover, hypoxia-induced exosomal miR-23a was also shown to inhibit the tight junction protein ZO-1 and increase vascular permeability[92]. It is noteworthy that most studies in this research area were conducted under acute hypoxic conditions. Umezu et al. were the first to report intercellular communication via exosome under chronic hypoxia[93]. A few hypoxia-resistant multiple myeloma (MM) cell lines were developed after incubation in hypoxic conditions for more than six months to mimic the hypoxic bone marrow environment in vivo[93]. Increased exosomal level of miR-135b was detected in these hypoxia-resistant MM cells, and it was shown to promote endothelial tube formation under hypoxia via the HIF-FIH signaling pathway[93].
Promotion of cancer cell invasion and metastasis by hypoxic tumor-derived exosomes
Exosomal ncRNAs within the hypoxic TME are known to regulate tumor invasion and metastasis by modulating EMT. In breast cancer, higher expression of miR-25-3p in hypoxic cancer-derived exosomes was found to stimulate cancer proliferation and migration by inducing IL-6 secretion and activating NF-κB signaling in macrophages[94]. In bladder cancer, the lncRNA (UCA1) preferentially secreted by hypoxic cancer cells was shown to promote cancer growth by stimulating EMT both in vitro and in vivo[95]. In oral squamous cell carcinoma, a higher level of miR-21 was detected in the exosomes from hypoxic cancer than those from normoxic cancer to promote migration and invasion by inducing EMT[96]. The interaction between stromal and cancer cells via exosome within the TME plays a critical role in the initiation of metastasis. Lung cancer cells have been shown to take up exosomes secreted from hypoxic bone marrow-derived mesenchymal stem cells (BMSCs) and acquire a greater tendency for invasion[97]. Three miRNAs (miR-193-3p, miR-210-3p, and miR-5100) showing a high abundance in hypoxic BMSC-derived exosomes were transferred to cancer cells and subsequently activated STAT3 signaling to induce EMT in the lung cancer cells[97]. Interestingly, these three miRNAs were also found to be upregulated in plasma-derived exosomes from lung cancer patients with metastatic disease than in non-metastatic patients[97].
Modulation of cancer immune system by hypoxic tumor-derived exosomes
Reduced immune surveillance is the major reason allowing primary tumors to develop metastasis in distant secondary organs[98,99]. Tumor-secreted exosomes have been reported to induce T-cell apoptosis, inhibit interferon gamma-dependent expression of macrophages, suppress natural killer (NK) cell activity, and increase myeloid-derived suppressor cell (MSDCs) population, which collectively suppress immune surveillance and allow tumor growth[100,101]. Figure 2 illustrates the major mechanisms by which hypoxic tumor-derived exosomes promote an immunosuppressive TME.
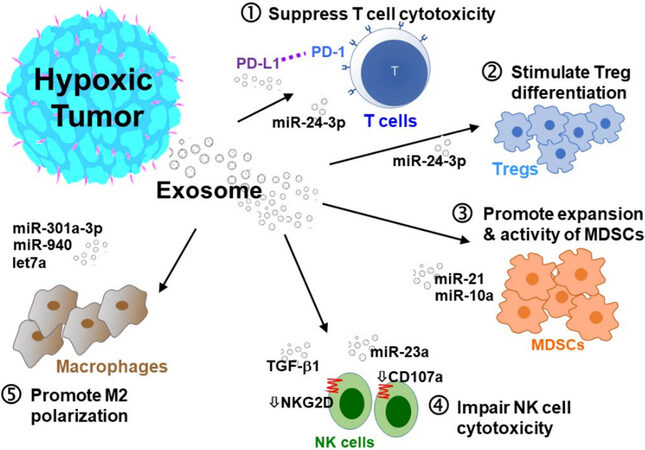
Figure 2. Hypoxic tumor-secreted exosomes promote an immunosuppressive TME. Hypoxic tumor-secreted exosomes promote an immunosuppressive TME by interfering with several intracellular pathways and modulating immune accessory cells, including cytotoxic T cells, T-regulatory cells (Tregs), myeloid-derived suppressor cells (MDSCs), natural killer cells (NK), and tumor-associated macrophages (TAMs). (1) Inhibition of T cell proliferation; (2) stimulation of Treg differentiation; (3) induction of MDSCs; (4) impairment of NK cells; and (5) stimulation of M2 polarization of TAMs.
Inhibition of T cell proliferation
Tumoral exosomes loaded with biologically active cargoes have been proposed to mediate the intercellular transmission of signals within the TME to promote immune escape and tumor progression. Ye et al. were the first to report a differential miRNA signature from nasopharyngeal carcinoma-derived exosomes to mediate T cell dysfunction[102]. The induction of exosomal miR-24-3p in nasopharyngeal carcinoma-derived exosomes under hypoxia was found to inhibit T cell proliferation but promote differentiation of T-regulatory cells (Tregs) by targeting FGF11 via the upregulation of p-ERK, p-STAT1, and p-STAT3 and downregulation of p-STAT5[103].
Immune checkpoint inhibitors including anti-programmed cell death receptor (PD-1) (nivolumab and pembrolizumab) or anti-PD-ligand (PD-L1) (duralumab, atezolizumab, and avelumab) monoclonal antibodies are revolutionizing cancer therapy. They lead to durable anticancer responses and overall survival benefits in a wide range of cancer types[104]. PD-1 is an inhibitory receptor expressed on activated T cells, B cells, and natural killer cells, which blunt the immune response under physiological conditions. The T cell-mediated cancer-killing effect will be suppressed when PD-1 is occupied by its major ligand PD-L1 (expressed in tumor cells and infiltrating immune cells). Anti-PD-1/PD-L1 antibodies work by binding to the inhibitory PD-1 receptors on tumor-reactive T cells and PD-L1 on tumor cells, respectively, to disrupt the PD-1/PD-L1 interaction and reactivate the cytotoxic T cell activity.
Despite the breakthrough of anti-PD-1/PD-L1 immunotherapy, the response rate is low. Moreover, most patients who initially respond to immunotherapy will eventually relapse because of adaptive resistance. To maximize the full potential of anti-PD-1/PD-L1 immunotherapy, the mechanisms underlying these de novo and adaptive resistance mechanisms is a research area of intensive investigation. When T cells recognize the tumor antigen on the cancer surface, they release interferons to induce PD-L1 expression in cancer cells[105]. The increased PD-L1 expression in cancer cells will then lead to specific inhibition of T cell recognition of cancer, subsequently resulting in a phenomenon known as adaptive immune resistance and inhibiting the antitumor immune response. To this end, PD-L1 loaded in exosomes was shown to interact directly with T cells to suppress anticancer efficacy of chemotherapy in various cancer types, including breast[106], gastric[107] head and neck[108], melanoma[109], pancreatic[110], and prostate[111] cancer. It will be useful to elucidate whether hypoxia regulates the loading of PD-L1 into tumor-derived exosomes.
Induction of MDSCs
MDSCs are a heterogeneous population of immune cells from the myeloid lineage which migrate to tumor sites to create an immunosuppressive TME[112]. MDSCs suppress adaptive and innate immunity by inhibiting T cell activation, promoting macrophage M2 polarization, inducing CAF differentiation, and inhibiting NK cell cytotoxicity. The abundance of MDSCs at tumor sites is known to correlate closely with poor clinical prognosis and reduce the efficacy of immunotherapy in cancer patients. In glioma, miR-21 and miR-10a secreted in tumor-derived exosomes under hypoxia have been reported to promote the expansion and activity of MDSCs in vitro and in vivo via the miR-21/PTEN/PI3K/AKT and miR-10a/RORA/IkBα/NF-κB pathways, respectively[113]. Therefore, novel strategies to modulate hypoxic tumor-secreted exosomes may be developed to regulate MDSCs and potentiate immunotherapy[114]. To this end, miR-21 loaded in γδ T cell-secreted exosomes has been shown to abate the function of MDSCs by targeting PTEN in a PD-L1-dependent manner[115].
Impairment of natural killer cells
Hypoxia is also known to promote an immunosuppressive TME by attenuating cytotoxic T cell and Impairment of natural killer (NK) cell-mediated tumor cell lysis. Berchem et al. were the first to report the secretion of non-coding RNAs in exosomes from hypoxic lung cancer cells to impair NK cell cytotoxicity[116]. Under hypoxic conditions, higher miR-23a expression was observed in lung cancer cell-derived exosomes, which impaired NK cell cytotoxicity by targeting CD107a[116]. Moreover, the hypoxic tumor-derived exosomes were also shown to transfer TGF-β1 to NK cells, thereby reducing the cell surface expression of the activating receptor NKG2D and inhibiting NK cell cytotoxicity[116].
Stimulation of M2 polarization of tumor-associated macrophages
TAMs refer to the major tumor-infiltrating immune cells, which interact with the tumors and tumor-associated macrophages (TME) to regulate tumor immunity[117]. Macrophage polarization is the process by which macrophages adopt distinct functional phenotypes in response to environmental stimuli and signals. M1 macrophages are functionally pro-inflammatory and antimicrobial, whereas M2 macrophages are anti-inflammatory. M1 and M2 macrophages exhibit a high degree of plasticity and are converted into each other upon changes within the TME or anticancer therapies. Under hypoxic pressure, tumor-derived exosomes have been shown to induce M2 polarization in various cancer types. In pancreatic cancer, miR-301a-3p was highly expressed in hypoxic cancer cell-derived exosomes, and it was shown to promote macrophage M2 polarization by activating PTEN/PI3Kγ signaling pathway[118]. Coculture of pancreatic cancer cells with the hypoxic cancer-derived exosomes or miR-301a-3p-upregulated macrophages was shown to facilitate the epithelial-mesenchymal transition and lung metastasis[118]. In epithelial ovarian cancer (EOC), hypoxic tumor-derived exosomes were shown to express a high level of miR-940, and they stimulated M2 polarization of macrophages and promoted cancer proliferation and migration[119]. A differential miRNA expression signature was also identified in the EOC-derived exosome under hypoxia to promote M2 polarization. miR-21-3p, miR-125b-5p, and miR-181d-5p were induced by HIF-1α and HIF-2α in the exosomes under hypoxic conditions, which regulate SOCS4/5-STAT3 signaling to stimulate M2 polarization and a malignant TME[120]. In melanoma, let-7a was shown to be downregulated in hypoxic cancer cells but remarkably increased in the hypoxic cancer-derived exosomes[121]. The exosomes carrying let-7a were found to promote a metabolic shift towards enhanced mitochondrial oxidative phosphorylation in macrophages by suppressing insulin-Akt-mTOR signaling to enhance cancer progression[121].
SUMMARY
Rapidly expanding and hypoxic tumors exploit exosomes to communicate with both cancerous and non-cancerous cells in the TME to promote cancer survival and resist immune surveillance. Hypoxia has been shown to directly induce the production of exosomes, modulate the exosome cargo sorting process, and promote exosome uptake by recipient cells. Under low oxygen tension, cancer cells are primed to glycolytic metabolism, thus inducing an acidic TME to indirectly promote intracellular transport of the exosome. Numerous regulatory molecules are involved in the regulation of exosome biology under hypoxia. More studies are warranted to fully unravel the effect of hypoxia on exosome-mediated intercellular communication within the TME.
FUTURE PERSPECTIVES
Advances in precision oncology have led to the increasing application of tissue and liquid biopsy methods in clinical practice to facilitate treatment selection and monitoring of cancer progression. For the traditional method using tissue biopsy, limited tissue specimens are taken from the patients. They are not able to reflect the spatial and temporal heterogeneity of a primary tumor or between multiple potentially discordant metastatic lesions. In comparison with tumor tissue analysis, liquid biopsy is less invasive, and the samples can be obtained throughout disease progression. The liquid biopsy analytes include circulating tumor cells, circulating nucleic acids[122], extracellular vesicles[123], and other tumor-derived materials present in blood and other body fluids. Among various liquid biopsy analytes, exosomes are unique in the way that they contain not only DNA but also RNAs, ncRNAs, proteins, glycoconjugates, and lipids, thus making them more versatile biomarkers.
Currently, the exogenous hypoxic marker drug pimonidazole has been used to visualize hypoxic regions in histological sections of tumors in pathological research in vivo[124]. However, the method is invasive and involves the surgical removal of tumors for imaging. Therefore, the application of tumor-derived exosomes from biological fluid to reveal the presence of hypoxic tumors will be beneficial. A few exosome biomarkers have been shown to reflect the hypoxic status of tumors as well as the stage of tumor progression. Exosomes derived from hypoxic glioma cells were enriched with hypoxia-related mRNA and proteins (including caveolin 1, IL-8, MMPs, and PDGF)[34]. Importantly, patients presenting with high levels of these biomarkers were associated with worse survival[34]. In rectal cancer patients, low levels of miR-486-5p and miR-181a-5p but high levels of miR-30d-5p in the exosomes harvested from plasma samples are associated with hypoxic tumors and poor prognosis[125]. Indeed, exosomes have been used as diagnostic or prognostic tools for assessing hypoxic tumors in recent clinical trials [Table 3].
Representative clinical trials exploiting exosome biomarkers to assess hypoxic tumors
| Cancer type | ClinicalTrials.gov Identifier | Aim(s) relevant to exosome biology | Current status |
| Lung cancer | NCT04629079 (LungExoDETECT) | • To validate exosomal assays that are based on hypoxia detection as potential biomarkers for early detection • Compare exosomal analysis with the standard of care imaging | Recruiting; Started in October 2020 |
| Lung cancer | NCT04529915 | • Multicenter clinical research for early diagnosis of lung cancer using exosomes derived from blood plasma | Active; Not recruiting; Started in April 2020 |
| Colorectal cancer | NCT04394572 (EXOSCOL01) | • To identify new diagnostic protein markers (e.g., integrins and metalloproteases) for colorectal cancer in circulating tumor exosomes | Recruiting; Started in January 2021 |
| Clear cell renal carcinoma | NCT04053855 | • To analyze urinary exosomes as a liquid biopsy tool for early diagnosis of clear cell renal cell carcinoma | Recruiting; Started in August 2019 |
| Ovarian cancer | NCT03738319 (EOC-EXOSOME) | • To analyze the expression of miRNA and lncRNA from exosomes in blood samples by next-generation sequencing in patients with high grade serous ovarian cancer or benign gynecologic diseases | Recruiting; Started in November 2018 |
| Melanoma | NCT02310451 | • Pilot study to examine exosomes collected from the blood before and after BRAF inhibitor therapy in patients with advanced unresectable or metastatic BRAF mutation-positive melanoma • To develop an exosome-based theranostic tool for personalized care in melanoma patients | Recruiting; Started in 2016 |
As hypoxic tumors produce exosomes to promote tumorigenesis, the inhibition of exosome formation and secretion may be exploited as a novel strategy to suppress tumor development. In an excellent recent review, He et al. summarized the strategies for exosomal targeting and discussed the potential clinical applications[126]. Experimental reagents such as manumycin A and GW4869 were shown to inhibit exosome biogenesis and secretion from mammalian cells[127]. On the other hand, the Rab family of GTPases involved in exosome secretion can also be targeted to hinder exosome-mediated intercellular communication. For example, Rab5a is involved in the early step of exosome biogenesis, whereas Rab11, Rab27a, and Rab35 regulate the fusion of multivesicular bodies with the plasma membrane and exosome secretion[128]. Downregulation of Rab27a has been shown to inhibit exosome-dependent and -independent tumor cell growth[128]. Specific inhibition of sphingomyelinase (an enzyme catalyzing the formation of ceramide from sphingomyelin) has also been shown to suppress exosome biogenesis and cargo loading, thereby retarding tumor growth[63].
The removal of oncogenic exosomes has been investigated as a novel therapeutic strategy for cancer therapy[129]. Mesoporous silica nanoparticles loaded with EGFR-targeting aptamers have been used to mop up circulating cancer-secreted EGFR+ exosomes, thus preventing their entry into the small intestine to suppress metastasis of lung cancer cells[129]. On the other hand, exosomes from immune cells were shown to exhibit anticancer activity[17]. Recently, Jiang et al. reported the induction of exosome production from NK cells under hypoxia[130]. More importantly, compared to normoxic conditions, NK cell-derived exosomes were found to express remarkably higher levels of FasL, perforin, and granzyme B in hypoxia to produce a higher NK cell cytotoxic effect[130]. Therefore, hypoxia-treated NK cells may be used to potentiate cancer immunotherapy.
Exosomes may also be employed as a drug delivery system for cancer therapy. Drugs or therapeutic siRNAs could be loaded into exosomes by various methods such as direct incubation, electroporation, and sonication[131]. Zhuang et al. reported the encapsulation of curcumin or an investigational STAT3 inhibitor (JSI124) in cancer cell-derived exosomes by incubating the exosomes with the drugs[132]. The drug-loaded exosomes were delivered to the brain for the treatment of inflammation via an intranasal route[132]. Besides, therapeutic siRNAs could also be loaded into the hydrophilic core of exosomes in the pharmaceutically active form[133,134]. Recently, Alvarez-Erviti et al. reported the successful delivery of siRNAs to the mouse brain using dendritic cell-derived exosomes[135]. Alternatively, the anticancer drug paclitaxel has been loaded indirectly into exosomes secreted from gingival mesenchymal stromal cells (MSCs) after co-culturing them with the drug[136]. Importantly, the exosomes derived from MSCs after priming with paclitaxel were shown to exhibit significant anticancer activity against human pancreatic cancer cells in vitro[136]. Interestingly, in a recent study examining anticancer drug delivery by exosomes, exosomes from hypoxic human breast cancer cells loaded with olaparib (a PARP inhibitor) were found to exhibit a superior uptake rate when they were co-cultured with hypoxic cancer cells[20]. A more detailed investigation of exosome loading and production under hypoxic conditions is advocated to further optimize exosome-mediated drug delivery.
DECLARATIONS
AcknowledgmentsWe acknowledge the researchers who have contributed to the understanding of tumor microenvironment regulation by cancer-derived exosomes in hypoxia and whose works have not been cited here because of space limitations.
Authors’ contributionsConception and preparation of this manuscript: To KKW, Cho WCS
Availability of data and materialsNot applicable.
Financial support and sponsorshipResearch in the To’s Lab was supported in part by research grants from the Chinese University of Hong Kong (Direct Grant 2019.084 & 2021.010).
Conflicts of interestAll authors declared that there are no conflicts of interest.
Ethical approval and consent to participateNot applicable.
Consent for publicationNot applicable.
Copyright© The Author(s) 2022.
REFERENCES
1. Belisario DC, Kopecka J, Pasino M, et al. Hypoxia dictates metabolic rewiring of tumors: implications for chemoresistance. Cells 2020;9:2598.
2. Kopecka J, Salaroglio IC, Perez-Ruiz E, et al. Hypoxia as a driver of resistance to immunotherapy. Drug Resist Updat 2021;59:100787.
3. Shamis SAK, McMillan DC, Edwards J. The relationship between hypoxia-inducible factor 1α (HIF-1α) and patient survival in breast cancer: Systematic review and meta-analysis. Crit Rev Oncol Hematol 2021;159:103231.
4. Jadli AS, Ballasy N, Edalat P, Patel VB. Inside(sight) of tiny communicator: exosome biogenesis, secretion, and uptake. Mol Cell Biochem 2020;467:77-94.
5. Braicu C, Tomuleasa C, Monroig P, Cucuianu A, Berindan-Neagoe I, Calin GA. Exosomes as divine messengers: are they the Hermes of modern molecular oncology? Cell Death Differ 2015;22:34-45.
6. Mathivanan S, Ji H, Simpson RJ. Exosomes: extracellular organelles important in intercellular communication. J Proteomics 2010;73:1907-20.
7. Samanta S, Rajasingh S, Drosos N, Zhou Z, Dawn B, Rajasingh J. Exosomes: new molecular targets of diseases. Acta Pharmacol Sin 2018;39:501-13.
8. Azmi AS, Bao B, Sarkar FH. Exosomes in cancer development, metastasis, and drug resistance: a comprehensive review. Cancer Metastasis Rev 2013;32:623-42.
9. Santos P, Almeida F. Role of Exosomal miRNAs and the tumor microenvironment in drug resistance. Cells 2020;9:1450.
10. Rai A, Greening DW, Chen M, Xu R, Ji H, Simpson RJ. Exosomes derived from human primary and metastatic colorectal cancer cells contribute to functional heterogeneity of activated fibroblasts by reprogramming their proteome. Proteomics 2019;19:e1800148.
11. Yekula A, Yekula A, Muralidharan K, Kang K, Carter BS, Balaj L. Extracellular vesicles in glioblastoma tumor microenvironment. Front Immunol 2019;10:3137.
12. He G, Peng X, Wei S, et al. Exosomes in the hypoxic TME: from release, uptake and biofunctions to clinical applications. Mol Cancer 2022;21:19.
13. Laplane L, Duluc D, Larmonier N, Pradeu T, Bikfalvi A. The multiple layers of the tumor environment. Trends Cancer 2018;4:802-9.
14. Godet I, Doctorman S, Wu F, Gilkes DM. Detection of hypoxia in cancer models: significance, challenges, and advances. Cells 2022;11:686.
15. Bosco MC, D'Orazi G, Del Bufalo D. Targeting hypoxia in tumor: a new promising therapeutic strategy. J Exp Clin Cancer Res 2020;39:8.
16. Niel G, D'Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol 2018;19:213-28.
17. Möller A, Lobb RJ. The evolving translational potential of small extracellular vesicles in cancer. Nat Rev Cancer 2020;20:697-709.
19. Wang Y, Yin K, Tian J, et al. Granulocytic myeloid-derived suppressor cells promote the stemness of colorectal cancer cells through exosomal S100A9. Adv Sci (Weinh) 2019;6:1901278.
20. Jung KO, Jo H, Yu JH, Gambhir SS, Pratx G. Development and MPI tracking of novel hypoxia-targeted theranostic exosomes. Biomaterials 2018;177:139-48.
21. Ren R, Sun H, Ma C, Liu J, Wang H. Colon cancer cells secrete exosomes to promote self-proliferation by shortening mitosis duration and activation of STAT3 in a hypoxic environment. Cell Biosci 2019;9:62.
22. Fu M, Gu J, Jiang P, Qian H, Xu W, Zhang X. Exosomes in gastric cancer: roles, mechanisms, and applications. Mol Cancer 2019;18:41.
23. Li J, Yuan H, Xu H, Zhao H, Xiong N. Hypoxic cancer-secreted exosomal miR-182-5p promotes glioblastoma angiogenesis by targeting kruppel-like factor 2 and 4. Mol Cancer Res 2020;18:1218-31.
24. Matsuura Y, Wada H, Eguchi H, et al. Exosomal miR-155 derived from hepatocellular carcinoma cells under hypoxia promotes angiogenesis in endothelial cells. Dig Dis Sci 2019;64:792-802.
25. Patton MC, Zubair H, Khan MA, Singh S, Singh AP. Hypoxia alters the release and size distribution of extracellular vesicles in pancreatic cancer cells to support their adaptive survival. J Cell Biochem 2020;121:828-39.
26. Liu W, Li L, Rong Y, et al. Hypoxic mesenchymal stem cell-derived exosomes promote bone fracture healing by the transfer of miR-126. Acta Biomater 2020;103:196-212.
27. Zhang W, Zhou X, Yao Q, Liu Y, Zhang H, Dong Z. HIF-1-mediated production of exosomes during hypoxia is protective in renal tubular cells. Am J Physiol Renal Physiol 2017;313:F906-13.
28. Zhu LP, Tian T, Wang JY, et al. Hypoxia-elicited mesenchymal stem cell-derived exosomes facilitates cardiac repair through miR-125b-mediated prevention of cell death in myocardial infarction. Theranostics 2018;8:6163-77.
29. Ostrowski M, Carmo NB, Krumeich S, et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat Cell Biol 2010;12:19-30; sup pp 1.
30. Rocha N, Kuijl C, van der Kant R, et al. Cholesterol sensor ORP1L contacts the ER protein VAP to control Rab7-RILP-p150 Glued and late endosome positioning. J Cell Biol 2009;185:1209-25.
31. Dorayappan KDP, Wanner R, Wallbillich JJ, et al. Hypoxia-induced exosomes contribute to a more aggressive and chemoresistant ovarian cancer phenotype: a novel mechanism linking STAT3/Rab proteins. Oncogene 2018;37:3806-21.
32. Zhang H, Freitas D, Kim HS, et al. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat Cell Biol 2018;20:332-43.
33. Willms E, Cabañas C, Mäger I, Wood MJA, Vader P. Extracellular vesicle heterogeneity: subpopulations, isolation techniques, and diverse functions in cancer progression. Front Immunol 2018;9:738.
34. Kucharzewska P, Christianson HC, Welch JE, et al. Exosomes reflect the hypoxic status of glioma cells and mediate hypoxia-dependent activation of vascular cells during tumor development. Proc Natl Acad Sci U S A 2013;110:7312-7.
36. Kalluri R, LeBleu VS. function
37. Navarro A, Molins L, Marrades RM, et al. Exosome analysis in tumor-draining pulmonary vein identifies NSCLC patients with higher risk of relapse after curative surgery. Cancers (Basel) 2019;11:249.
38. Ramteke A, Ting H, Agarwal C, et al. Exosomes secreted under hypoxia enhance invasiveness and stemness of prostate cancer cells by targeting adherens junction molecules: hypoxic-exosomes role in pca aggressiveness. Mol Carcinog 2015;54:554-65.
39. Caponnetto F, Manini I, Skrap M, et al. Size-dependent cellular uptake of exosomes. Nanomedicine 2017;13:1011-20.
40. Kumar A, Deep G. Exosomes in hypoxia-induced remodeling of the tumor microenvironment. Cancer Letters 2020;488:1-8.
41. Wang W, Han Y, Jo HA, Lee J, Song YS. Non-coding RNAs shuttled via exosomes reshape the hypoxic tumor microenvironment. J Hematol Oncol 2020;13:67.
42. Yang W, Ma J, Zhou W, Cao B, Zhou X, Zhang H, et al. Reciprocal regulations between miRNAs and HIF-1a in human cancers. Cell Mol Life Sci 2019; 76:453-71.
43. Bruning U, Cerone L, Neufeld Z, et al. MicroRNA-155 promotes resolution of hypoxia-inducible factor 1alpha activity during prolonged hypoxia. Mol Cell Biol 2011;31:4087-96.
44. Fabbiano F, Corsi J, Gurrieri E, Trevisan C, Notarangelo M, D’Agostino VG. RNA packaging into extracellular vesicles: an orchestra of RNA-binding proteins? J Extracell Vesicles 2020;10:e12043.
45. Santangelo L, Giurato G, Cicchini C, et al. The RNA-binding protein SYNCRIP is a component of the hepatocyte exosomal machinery controlling microRNA sorting. Cell Rep 2016;17:799-808.
46. Han J, Li J, Ho JC, et al. Hypoxia is a key driver of alternative splicing in human breast cancer cells. Sci Rep 2017;7:4108.
47. Farina AR, Cappabianca L, Sebastiano M, Zelli V, Guadagni S, Mackay AR. Hypoxia-induced alternative splicing: the 11th Hallmark of cancer. J Exp Clin Cancer Res 2020;39:110.
48. Nigita G, Acunzo M, Romano G, et al. microRNA editing in seed region aligns with cellular changes in hypoxic conditions. Nucleic Acids Res 2016;44:6298-308.
49. Lin F, Zeng Z, Song Y, et al. YBX-1 mediated sorting of miR-133 into hypoxia/reoxygenation-induced EPC-derived exosomes to increase fibroblast angiogenesis and MEndoT. Stem Cell Res Ther 2019;10:263.
50. Qiu W, Guo X, Li B, et al. Exosomal miR-1246 from glioma patient body fluids drives the differentiation and activation of myeloid-derived suppressor cells. Mol Ther 2021;29:3449-64.
51. Nallamshetty S, Chan SY, Loscalzo J. Hypoxia: a master regulator of microRNA biogenesis and activity. Free Radic Biol Med 2013;64:20-30.
53. Xia X, Wang S, Ni B, et al. Hypoxic gastric cancer-derived exosomes promote progression and metastasis via MiR-301a-3p/PHD3/HIF-1α positive feedback loop. Oncogene 2020;39:6231-44.
54. Zeng Z, Zhao Y, Chen Q, et al. Hypoxic exosomal HIF-1α-stabilizing circZNF91 promotes chemoresistance of normoxic pancreatic cancer cells via enhancing glycolysis. Oncogene 2021;40:5505-17.
55. Jeppesen DK, Fenix AM, Franklin JL, et al. Reassessment of exosome composition. Cell 2019;177:428-445.e18.
56. Moreno-Gonzalo O, Villarroya-Beltri C, Sánchez-Madrid F. Post-translational modifications of exosomal proteins. Front Immunol 2014;5:383.
57. Wade BE, Zhao J, Ma J, Hart CM, Sutliff RL. Hypoxia-induced alterations in the lung ubiquitin proteasome system during pulmonary hypertension pathogenesis. Pulm Circ 2018;8:2045894018788267.
58. Sato A, Rahman NIA, Shimizu A, Ogita H. Cell-to-cell contact-mediated regulation of tumor behavior in the tumor microenvironment. Cancer Sci 2021;112:4005-12.
59. Martins ÁM, Ramos CC, Freitas D, Reis CA. Glycosylation of cancer extracellular vesicles: capture strategies, functional roles and potential clinical applications. Cells 2021;10:109.
60. Weinhouse GL, Belloni PN, Farber HW. Effect of hypoxia on endothelial cell surface glycoprotein expression: modulation of glycoprotein IIIa and other specific surface glycoproteins. Exp Cell Res 1993;208:465-78.
61. Cerezo-magaña M, Christianson HC, van Kuppevelt TH, Forsberg-nilsson K, Belting M. Hypoxic induction of exosome uptake through proteoglycan-dependent endocytosis fuels the lipid droplet phenotype in glioma. Molecular Cancer Research 2021;19:528-40.
62. Hannafon BN, Gin AL, Xu YF, Bruns M, Calloway CL, Ding WQ. Metastasis-associated protein 1 (MTA1) is transferred by exosomes and contributes to the regulation of hypoxia and estrogen signaling in breast cancer cells. Cell Commun Signal 2019;17:13.
63. Trajkovic K, Hsu C, Chiantia S, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008;319:1244-7.
64. Kajimoto T, Okada T, Miya S, Zhang L, Nakamura S. Ongoing activation of sphingosine 1-phosphate receptors mediates maturation of exosomal multivesicular endosomes. Nat Commun 2013;4:2712.
65. Namba Y, Sogawa C, Okusha Y, et al. Depletion of lipid efflux pump ABCG1 triggers the intracellular accumulation of extracellular vesicles and reduces aggregation and tumorigenesis of metastatic cancer cells. Front Oncol 2018;8:376.
66. Wei X, Liu C, Wang H, et al. Surface phosphatidylserine is responsible for the internalization on microvesicles derived from hypoxia-induced human bone marrow mesenchymal stem cells into human endothelial cells. PLoS One 2016;11:e0147360.
67. Schlaepfer IR, Nambiar DK, Ramteke A, et al. Hypoxia induces triglycerides accumulation in prostate cancer cells and extracellular vesicles supporting growth and invasiveness following reoxygenation. Oncotarget 2015;6:22836-56.
68. Xia QS, Lu FE, Wu F, et al. New role for ceramide in hypoxia and insulin resistance. World J Gastroenterol 2020;26:2177-86.
69. Ban JJ, Lee M, Im W, Kim M. Low pH increases the yield of exosome isolation. Biochem Biophys Res Commun 2015;461:76-9.
70. Gong C, Zhang X, Shi M, et al. Tumor exosomes reprogrammed by low pH are efficient targeting vehicles for smart drug delivery and personalized therapy against their homologous tumor. Adv Sci (Weinh) 2021;8:2002787.
71. Wang Y, Roche O, Xu C, et al. Hypoxia promotes ligand-independent EGF receptor signaling via hypoxia-inducible factor-mediated upregulation of caveolin-1. Proc Natl Acad Sci USA 2012;109:4892-7.
72. Mulcahy LA, Pink RC, Carter DR. Routes and mechanisms of extracellular vesicle uptake. J Extracell Vesicles 2014;3:24641.
73. Dang K, Myers KA. The role of hypoxia-induced miR-210 in cancer progression. Int J Mol Sci 2015;16:6353-72.
74. King HW, Michael MZ, Gleadle JM. Hypoxic enhancement of exosome release by breast cancer cells. BMC Cancer 2012;12:421.
75. Wozniak M, Peczek L, Czernek L, Düchler M. Analysis of the miRNA profiles of melanoma exosomes derived under normoxic and hypoxic culture conditions. Anticancer Res 2017;37:6779-89.
76. Sharma KD, Schaal D, Kore RA, et al. Glioma-derived exosomes drive the differentiation of neural stem cells to astrocytes. PLoS One 2020;15:e0234614.
77. Dong C, Liu X, Wang H, et al. Hypoxic non-small-cell lung cancer cell-derived exosomal miR-21 promotes resistance of normoxic cell to cisplatin. Onco Targets Ther 2019;12:1947-56.
78. Takahashi K, Yan IK, Kogure T, Haga H, Patel T. Extracellular vesicle-mediated transfer of long non-coding RNA ROR modulates chemosensitivity in human hepatocellular cancer. FEBS Open Bio 2014;4:458-67.
79. Zhu X, Shen H, Yin X, et al. Macrophages derived exosomes deliver miR-223 to epithelial ovarian cancer cells to elicit a chemoresistant phenotype. J Exp Clin Cancer Res 2019;38:81.
80. Bebawy M, Combes V, Lee E, et al. Membrane microparticles mediate transfer of P-glycoprotein to drug sensitive cancer cells. Leukemia 2009;23:1643-9.
81. Corcoran C, Rani S, O'Brien K, et al. Docetaxel-resistance in prostate cancer: evaluating associated phenotypic changes and potential for resistance transfer via exosomes. PLoS One 2012;7:e50999.
82. Wei Y, Lai X, Yu S, et al. Exosomal miR-221/222 enhances tamoxifen resistance in recipient ER-positive breast cancer cells. Breast Cancer Res Treat 2014;147:423-31.
83. Zhang FF, Zhu YF, Zhao QN, et al. Microvesicles mediate transfer of P-glycoprotein to paclitaxel-sensitive A2780 human ovarian cancer cells, conferring paclitaxel-resistance. Eur J Pharmacol 2014;738:83-90.
84. Sousa D, Lima RT, Vasconcelos MH. Intercellular transfer of cancer drug resistance traits by extracellular vesicles. Trends Mol Med 2015;21:595-608.
85. Lv MM, Zhu XY, Chen WX, et al. Exosomes mediate drug resistance transfer in MCF-7 breast cancer cells and a probable mechanism is delivery of P-glycoprotein. Tumour Biol 2014;35:10773-9.
86. Kato T, Mizutani K, Kameyama K, et al. Serum exosomal P-glycoprotein is a potential marker to diagnose docetaxel resistance and select a taxoid for patients with prostate cancer. Urol Oncol 2015;33:385.e15-20.
87. Torreggiani E, Roncuzzi L, Perut F, Zini N, Baldini N. Multimodal transfer of MDR by exosomes in human osteosarcoma. Int J Oncol 2016;49:189-96.
88. Safaei R, Larson BJ, Cheng TC, et al. Abnormal lysosomal trafficking and enhanced exosomal export of cisplatin in drug-resistant human ovarian carcinoma cells. Mol Cancer Ther 2005;4:1595-604.
89. Ciravolo V, Huber V, Ghedini GC, et al. Potential role of HER2-overexpressing exosomes in countering trastuzumab-based therapy. J Cell Physiol 2012;227:658-67.
90. Schito L. Hypoxia-dependent angiogenesis and lymphangiogenesis in cancer. In: Gilkes DM, editor. Hypoxia and cancer metastasis. Cham: Springer International Publishing; 2019. pp. 71-85.
91. Guo Z, Wang X, Yang Y, et al. Hypoxic tumor-derived exosomal long noncoding RNA UCA1 promotes angiogenesis via miR-96-5p/AMOTL2 in pancreatic cancer. Mol Ther Nucleic Acids 2020;22:179-95.
92. Hsu YL, Hung JY, Chang WA, et al. Hypoxic lung cancer-secreted exosomal miR-23a increased angiogenesis and vascular permeability by targeting prolyl hydroxylase and tight junction protein ZO-1. Oncogene 2017;36:4929-42.
93. Umezu T, Tadokoro H, Azuma K, Yoshizawa S, Ohyashiki K, Ohyashiki JH. Exosomal miR-135b shed from hypoxic multiple myeloma cells enhances angiogenesis by targeting factor-inhibiting HIF-1. Blood 2014;124:3748-57.
94. Fisher L. Retraction: Exosomal miR-25-3p derived from hypoxia tumor mediates IL-6 secretion and stimulates cell viability and migration in breast cancer. RSC Adv 2021;11:6257.
95. Xue M, Chen W, Xiang A, et al. Hypoxic exosomes facilitate bladder tumor growth and development through transferring long non-coding RNA-UCA1. Mol Cancer 2017;16:143.
96. Li L, Li C, Wang S, et al. Exosomes derived from hypoxic oral squamous cell carcinoma cells deliver miR-21 to normoxic cells to elicit a prometastatic phenotype. Cancer Res 2016;76:1770-80.
97. Zhang X, Sai B, Wang F, et al. Hypoxic BMSC-derived exosomal miRNAs promote metastasis of lung cancer cells via STAT3-induced EMT. Mol Cancer 2019;18:40.
98. Liu Y, Cao X. Characteristics and significance of the pre-metastatic niche. Cancer Cell 2016;30:668-81.
99. Cole K, Pravoverov K, Talmadge JE. Role of myeloid-derived suppressor cells in metastasis. Cancer Metastasis Rev 2021;40:391-411.
100. Arkhypov I, Lasser S, Petrova V, et al. Myeloid cell modulation by tumor-derived extracellular vesicles. Int J Mol Sci 2020;21:6319.
101. Olejarz W, Dominiak A, Żołnierzak A, Kubiak-Tomaszewska G, Lorenc T. Tumor-derived exosomes in immunosuppression and immunotherapy. J Immunol Res 2020;2020:6272498.
102. Agnoletto C, Melloni E, Casciano F, et al. Sodium dichloroacetate exhibits anti-leukemic activity in B-chronic lymphocytic leukemia (B-CLL) and synergizes with the p53 activator Nutlin-3. Oncotarget 2014;5:4347-60.
103. Ye SB, Zhang H, Cai TT, et al. Exosomal miR-24-3p impedes T-cell function by targeting FGF11 and serves as a potential prognostic biomarker for nasopharyngeal carcinoma. J Pathol 2016;240:329-40.
104. Wang J, Yang T, Xu J. Therapeutic development of immune checkpoint inhibitors. In: Xu J, editor. Regulation of cancer immune checkpoints. Singapore: Springer; 2020. pp. 619-49.
105. Garcia-Diaz A, Shin DS, Moreno BH, et al. Interferon receptor signaling pathways regulating PD-L1 and PD-L2 expression. Cell Rep 2017;19:1189-201.
106. Yang Y, Li CW, Chan LC, et al. Exosomal PD-L1 harbors active defense function to suppress T cell killing of breast cancer cells and promote tumor growth. Cell Res 2018;28:862-4.
107. Fan Y, Che X, Qu J, et al. Exosomal PD-L1 retains immunosuppressive activity and is associated with gastric cancer prognosis. Ann Surg Oncol 2019;26:3745-55.
108. Theodoraki MN, Yerneni SS, Hoffmann TK, Gooding WE, Whiteside TL. Clinical significance of PD-L1+ exosomes in plasma of head and neck cancer patients. Clin Cancer Res 2018;24:896-905.
109. Chen G, Huang AC, Zhang W, et al. Exosomal PD-L1 contributes to immunosuppression and is associated with anti-PD-1 response. Nature 2018;560:382-6.
110. Lux A, Kahlert C, Grützmann R, Pilarsky C. c-Met and PD-L1 on circulating exosomes as diagnostic and prognostic markers for pancreatic cancer. Int J Mol Sci 2019;20:3305.
111. Poggio M, Hu T, Pai CC, et al. Suppression of exosomal PD-L1 induces systemic anti-tumor immunity and memory. Cell 2019;177:414-427.e13.
112. Ostrand-Rosenberg S. Myeloid derived-suppressor cells: their role in cancer and obesity. Curr Opin Immunol 2018;51:68-75.
113. Guo X, Qiu W, Liu Q, et al. Immunosuppressive effects of hypoxia-induced glioma exosomes through myeloid-derived suppressor cells via the miR-10a/Rora and miR-21/Pten Pathways. Oncogene 2018;37:4239-59.
114. Messmer MN, Netherby CS, Banik D, Abrams SI. Tumor-induced myeloid dysfunction and its implications for cancer immunotherapy. Cancer Immunol Immunother 2015;64:1-13.
115. Li L, Cao B, Liang X, et al. Microenvironmental oxygen pressure orchestrates an anti- and pro-tumoral γδ T cell equilibrium via tumor-derived exosomes. Oncogene 2019;38:2830-43.
116. Berchem G, Noman MZ, Bosseler M, et al. Hypoxic tumor-derived microvesicles negatively regulate NK cell function by a mechanism involving TGF-β and miR23a transfer. Oncoimmunology 2016;5:e1062968.
117. Pan Y, Yu Y, Wang X, Zhang T. Tumor-associated macrophages in tumor immunity. Front Immunol 2020;11:583084.
118. Wang X, Luo G, Zhang K, et al. Hypoxic tumor-derived exosomal miR-301a mediates M2 macrophage polarization via PTEN/PI3Kγ to promote pancreatic cancer metastasis. Cancer Res 2018;78:4586-98.
119. Chen X, Ying X, Wang X, Wu X, Zhu Q, Wang X. Exosomes derived from hypoxic epithelial ovarian cancer deliver microRNA-940 to induce macrophage M2 polarization. Oncol Rep 2017;38:522-8.
120. Chen X, Zhou J, Li X, Wang X, Lin Y, Wang X. Exosomes derived from hypoxic epithelial ovarian cancer cells deliver microRNAs to macrophages and elicit a tumor-promoted phenotype. Cancer Lett 2018;435:80-91.
121. Park JE, Dutta B, Tse SW, et al. Hypoxia-induced tumor exosomes promote M2-like macrophage polarization of infiltrating myeloid cells and microRNA-mediated metabolic shift. Oncogene 2019;38:5158-73.
122. Alix-Panabières C, Pantel K. Clinical applications of circulating tumor cells and circulating tumor DNA as liquid biopsy. Cancer Discov 2016;6:479-91.
123. Yu W, Hurley J, Roberts D, et al. Exosome-based liquid biopsies in cancer: opportunities and challenges. Ann Oncol 2021;32:466-77.
124. Kizaka-Kondoh S, Konse-Nagasawa H. Significance of nitroimidazole compounds and hypoxia-inducible factor-1 for imaging tumor hypoxia. Cancer Sci 2009;100:1366-73.
125. Bjørnetrø T, Redalen KR, Meltzer S, et al. An experimental strategy unveiling exosomal microRNAs 486-5p, 181a-5p and 30d-5p from hypoxic tumour cells as circulating indicators of high-risk rectal cancer. J Extracell Vesicles 2019;8:1567219.
126. He J, Ren W, Wang W, et al. Exosomal targeting and its potential clinical application. Drug Deliv Transl Res 2021; doi: 10.1007/s13346-021-01087-1.
127. Zhou X, Zhang W, Yao Q, et al. Exosome production and its regulation of EGFR during wound healing in renal tubular cells. Am J Physiol Renal Physiol 2017;312:F963-70.
128. Bobrie A, Krumeich S, Reyal F, et al. Rab27a supports exosome-dependent and -independent mechanisms that modify the tumor microenvironment and can promote tumor progression. Cancer Res 2012;72:4920-30.
129. Xie X, Nie H, Zhou Y, et al. Eliminating blood oncogenic exosomes into the small intestine with aptamer-functionalized nanoparticles. Nat Commun 2019;10:5476.
130. Jiang Y, Jiang H, Wang K, Liu C, Man X, Fu Q. Hypoxia enhances the production and antitumor effect of exosomes derived from natural killer cells. Ann Transl Med 2021;9:473.
131. Bunggulawa EJ, Wang W, Yin T, et al. Recent advancements in the use of exosomes as drug delivery systems. J Nanobiotechnology 2018;16:81.
132. Zhuang X, Xiang X, Grizzle W, et al. Treatment of brain inflammatory diseases by delivering exosome encapsulated anti-inflammatory drugs from the nasal region to the brain. Mol Ther 2011;19:1769-79.
133. Aryani A, Denecke B. Exosomes as a nanodelivery system: a key to the future of neuromedicine? Mol Neurobiol 2016;53:818-34.
134. Jiang XC, Gao JQ. Exosomes as novel bio-carriers for gene and drug delivery. Int J Pharm 2017;521:167-75.
135. Alvarez-Erviti L, Seow Y, Yin H, Betts C, Lakhal S, Wood MJ. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol 2011;29:341-5.
136. Coccè V, Franzè S, Brini AT, et al. In vitro anticancer activity of extracellular vesicles (EVs) secreted by gingival mesenchymal stromal cells primed with paclitaxel. Pharmaceutics 2019;11:61.
137. Huang Z, Yang M, Li Y, Yang F, Feng Y. Exosomes derived from hypoxic colorectal cancer cells transfer Wnt4 to normoxic cells to elicit a prometastatic phenotype. Int J Biol Sci 2018;14:2094-102.
138. Kore RA, Edmondson JL, Jenkins SV, et al. Hypoxia-derived exosomes induce putative altered pathways in biosynthesis and ion regulatory channels in glioblastoma cells. Biochem Biophys Rep 2018;14:104-13.
139. Shan Y, You B, Shi S, et al. Hypoxia-induced matrix metalloproteinase-13 expression in exosomes from nasopharyngeal carcinoma enhances metastases. Cell Death Dis 2018;9:382.
140. Aga M, Bentz GL, Raffa S, et al. Exosomal HIF1α supports invasive potential of nasopharyngeal carcinoma-associated LMP1-positive exosomes. Oncogene 2014;33:4613-22.
Cite This Article
Export citation file: BibTeX | RIS
OAE Style
To KKW, Cho WCS. Exosome secretion from hypoxic cancer cells reshapes the tumor microenvironment and mediates drug resistance. Cancer Drug Resist 2022;5:577-94. http://dx.doi.org/10.20517/cdr.2022.38
AMA Style
To KKW, Cho WCS. Exosome secretion from hypoxic cancer cells reshapes the tumor microenvironment and mediates drug resistance. Cancer Drug Resistance. 2022; 5(3): 577-94. http://dx.doi.org/10.20517/cdr.2022.38
Chicago/Turabian Style
To, Kenneth K. W., William C. S. Cho. 2022. "Exosome secretion from hypoxic cancer cells reshapes the tumor microenvironment and mediates drug resistance" Cancer Drug Resistance. 5, no.3: 577-94. http://dx.doi.org/10.20517/cdr.2022.38
ACS Style
To, KKW.; Cho WCS. Exosome secretion from hypoxic cancer cells reshapes the tumor microenvironment and mediates drug resistance. Cancer Drug Resist. 2022, 5, 577-94. http://dx.doi.org/10.20517/cdr.2022.38
About This Article
Special Issue
Copyright

Data & Comments
Data


 Cite This Article 11 clicks
Cite This Article 11 clicks

 Like This Article 2
likes
Like This Article 2
likes








Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.